Block5: Lernprozesse
Aplysia, Synaptische Sensitisierung, Lang- und Kurzzeitgedächtnis, CREB1/2; Konditionierung, Langzeit-Potenzierung(LTP) und Langzeit-Depression(LTD)
Fragen:
- Welche Stadien des Gedächtnisses lassen sich unterscheiden?
- Welche Gedächtnissysteme kann man unterscheiden?
- Grundlagenforschung: Welche neuronalen Prozesse spielen beim Kurzzeit-Gedächtnis der Meeresschnecke Aplysia eine Rolle?
- Welche neuronalen Prozesse spielen beim Langzeit-Gedächtnis von Aplysia eine Rolle?
- Was könnten Regulator-Proteine CREB1/CREB2 des Zellkerns mit Supergedächtnisleistungen bzw. Gedächtnislücken zu tun haben?
- Bilden Prionen einen Schlüssel zur Erinnerungsfähigkeit?
- Wie lassen sich synaptische Übertragungseigenschaften modulieren?
- Assoziatives Lernen: Wie lässt sich klassische Konditionierung durch Hebb-Synapsen erklären?
- Welches Trainingsmuster kommt bei der klassischen Konditionierung zum Einsatz?
- Furcht-Konditionierung bei der Ratte durch Langzeitpotenzierung (LTP): Welche Neuronenschaltung könnte zugrundeliegen?
- Furcht-Konditionierung bei der Ratte: Auf welchen neurochemischen Prozessen könnte die LTP beruhen?
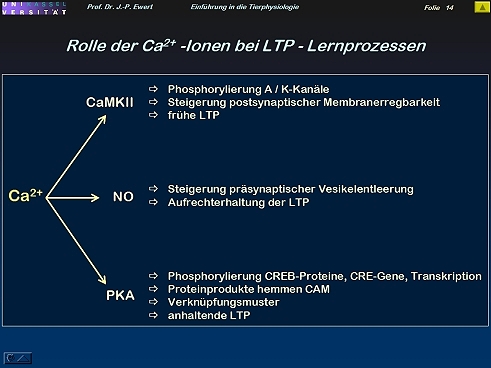
- Welche Wirkungen können Ca 2+ Ionen bei LTP entfalten?
- Lidschlag-Konditionierung beim Kaninchen durch Langzeitdepression (LTD): Wie wird Lidschlag-Konditionierung induziert?
- Lidschlag-Konditionierung beim Kaninchen durch LTD: Welche Neuronenschaltung könnte zugrundeliegen?
- Lidschlag-Konditionierung beim Kaninchen: Auf welchen neurochemischen Prozessen könnte die LTD beruhen?
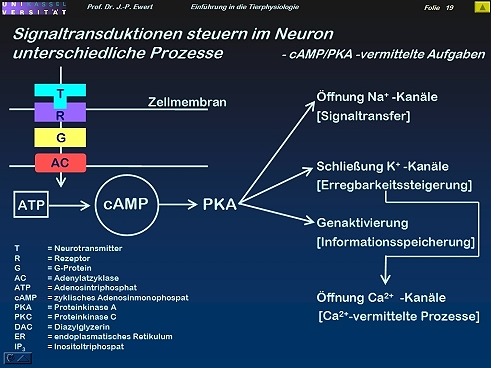
- Zusammenfassende Übersicht (I): Welche Prozesse im Neuron werden durch cAMP/PKA gesteuert?
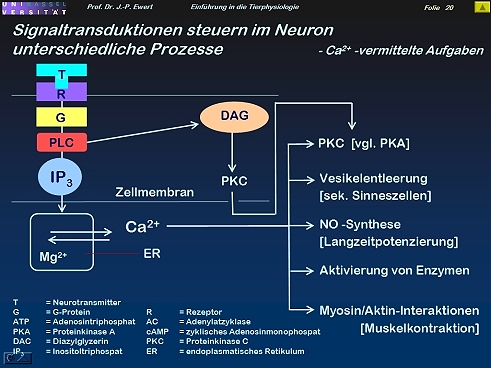
- Zusammenfassende Übersicht (II): Welche Prozesse im Neuron werden durch IP3/Ca 2+ gesteuert?
- Zusammenfassende Übersicht (III): Welche Folgeprozesse im Neuron werden durch den Mediator NO beeinflusst?
Folien:
Gedächtnisforschung im Spiegel der Medien - Eric Kandel vom Nobelpreis 2000 zur Entwicklung einer Gedächtnis-Pille: Nachrichten VOX Januar 2005; Gedächtnisbildung im Focus des Magazins Focus 47/2003; sind Erinnerungen ansteckend? FAZ 10.01.2004
Welche Stadien des Gedächtnisses lassen sich unterscheiden?
1

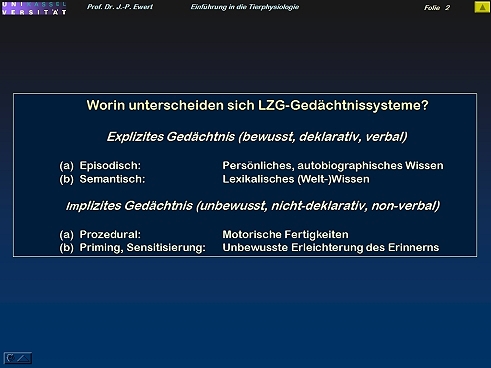
Welche Gedächtnissysteme kann man unterscheiden?
2
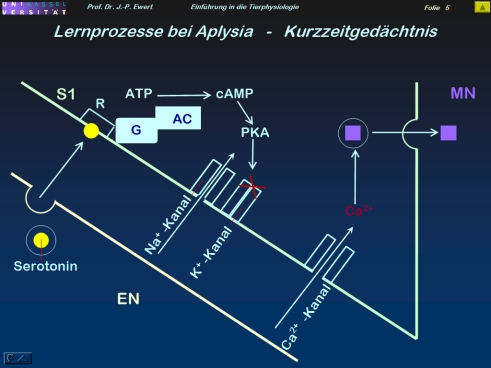
Grundlagenforschung: Welche neuronalen Prozesse spielen beim Kurzzeit-Gedächtnis der Meeresschnecke Aplysia eine Rolle?
3
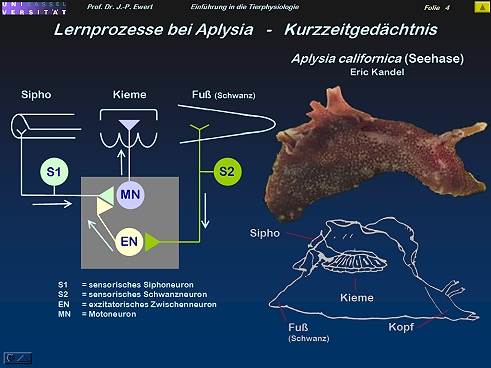
4
Welche neuronalen Prozesse spielen beim Langzeit-Gedächtnis der Meeresschnecke Aplysia eine Rolle?
5
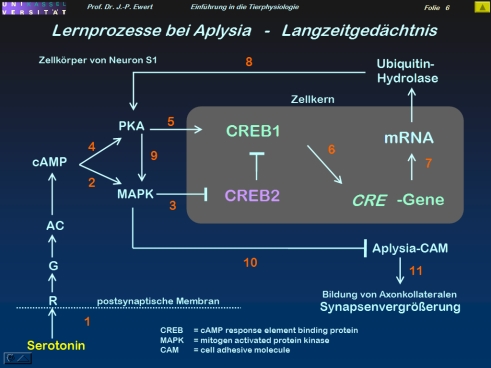
(3-5: Abstrahiert und modifiziert nach Kandel 1979; Abel et al. 1998)
Was könnten Regulator-Proteine CREB1/CREB2 des Zellkerns mit Supergedächtnisleistungen bzw. Gedächtnislücken zu tun haben?
6
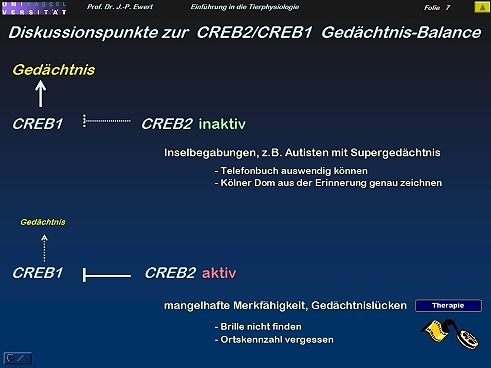
7

Bilden Prionen einen Schlüssel zur Erinnerungsfähigkeit?
8

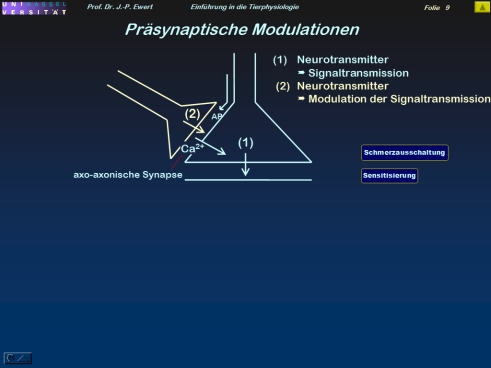
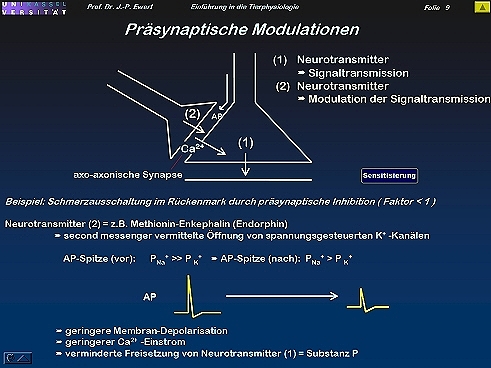
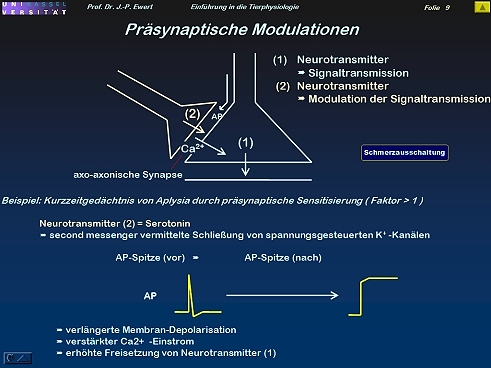
Wie lassen sich synaptische Übertragungseigenschaften modulieren?
9
Assoziatives Lernen: Wie lässt sich klassische Konditionierung durch Hebb-Synapsen erklären?
10
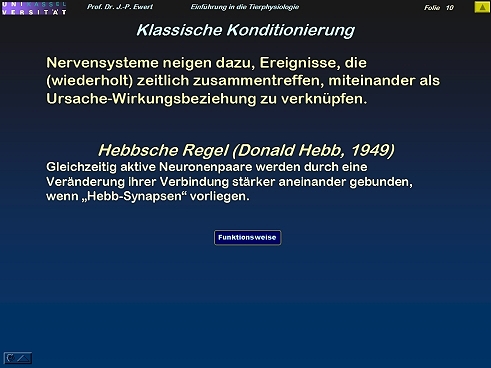

(Nach Hebb 1949)
Welches Trainingsmuster kommt bei der klassischen Konditionierung zum Einsatz?
11

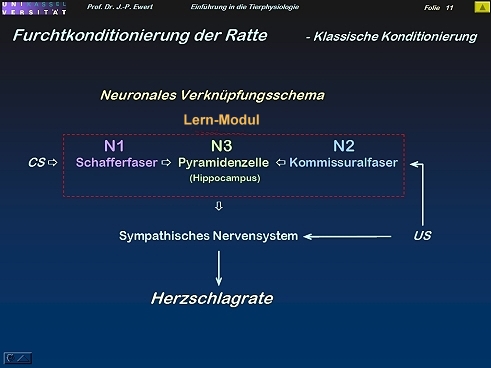
Furcht-Konditionierung bei der Ratte durch Langzeitpotenzierung (LTP): Welche Neuronenschaltung könnte zugrundeliegen?
12
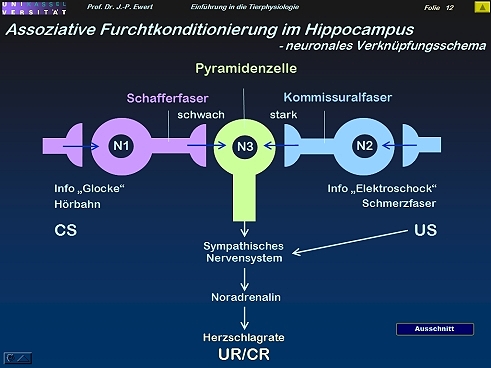
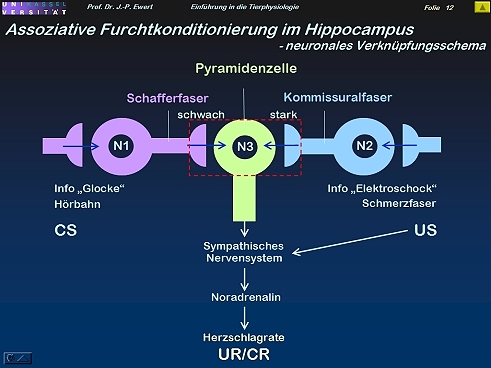
Furcht-Konditionierung bei der Ratte: Auf welchen neurochemischen Prozessen könnte die LTP beruhen?
13
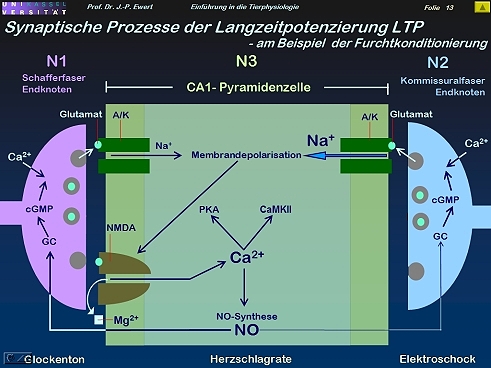
(Abstrahiert und modifiziert nach Kandel & Hawkins 1992; Hölscher 1997)
Welche Wirkungen können Ca 2+ Ionen bei LTP entfalten?
14
Lidschlag-Konditionierung beim Kaninchen durch Langzeitdepression (LTD): Wie wird diese Konditionierung induziert?
15
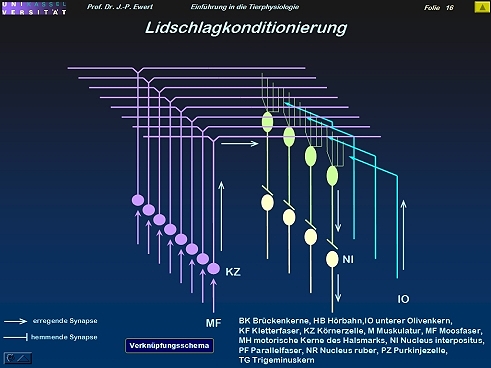
(Kombiniert nach Ewert 1980)
(Kombiniert und modifiziert nach Kim & Thompson 1997)
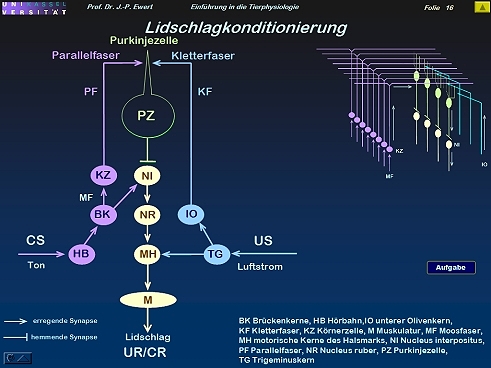
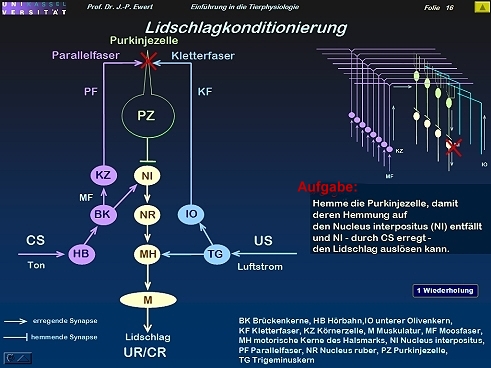
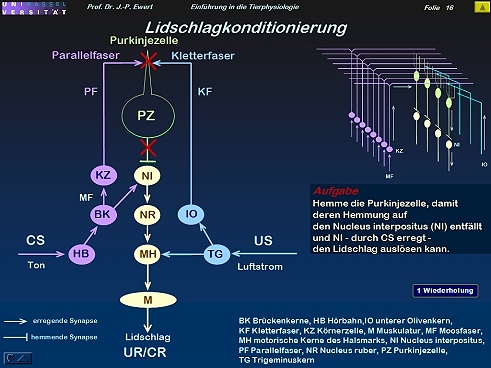
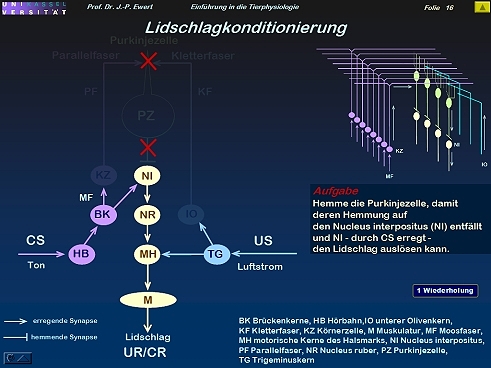
Lidschlag-Konditionierung beim Kaninchen durch LTD: Welche Neuronenschaltung könnte zugrundeliegen?
16
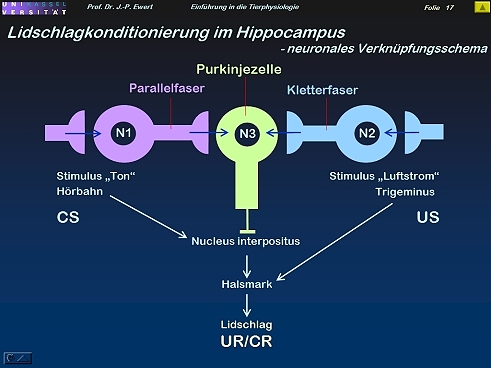
Lidschlag-Konditionierung beim Kaninchen: Auf welchen neurochemischen Prozessen könnte die LTD beruhen?
17
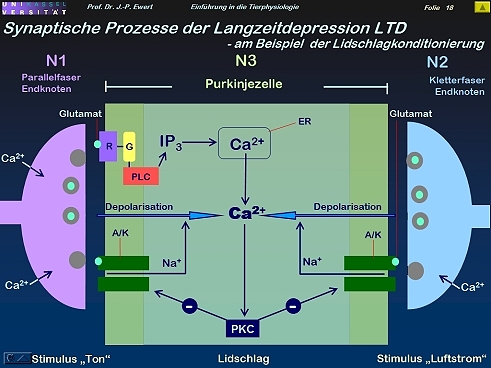
(Kombiniert und modifiziert nach Linden 1994)
Zusammenfassende Übersicht (I): Welche Prozesse im Neuron werden durch cAMP/PKA gesteuert?
18
Zusammenfassende Übersicht (II): Welche Prozesse im Neuron werden durch IP3/Ca 2+ gesteuert?
19
Zusammenfassende Übersicht (III): Welche Folgeprozesse im Neuron werden durch den Mediator NO beeinflusst?
20

